Одобрено Ученым советом Государственного комитета Совета Министров СССР по профессионально-техническому образованию в качестве учебного пособия для сельских профессионально-технических училищ и подготовки рабочих на производстве.
Москва "Колос" 1978
Введение
С расширением спортивных и эстетических интересов советских людей роль рысистой лошади как участницы увлекательнейшего спортивного зрелища возрастает. Соревнования наших наездников с зарубежными расширяют культурные контакты с другими странами. Рысистые лошади высокого резвостного класса пользуются большим спросом на внешнем рынке. Советское коннозаводство заинтересовано в том, чтобы работать с наиболее перспективными рысистыми лошадьми и чтобы применяемая к ним система тренировки была наиболее эффективной. Такой может быть лишь система, опирающаяся на понимание глубоких физиологических основ работоспособности рысистой лошади и с этих позиций творчески воспринимающая достижения передовой практики.
Двухвековая практика работы с рысистыми лошадьми создала арсенал методов и приемов их тренировки, большинство из которых широко применяется как в отечественном, так и в зарубежном коннозаводстве. Однако эмпирически найденные приемы тренировки не всегда оказываются верными и полезными, поскольку полезное для одной лошади в одних условиях может оказаться вредным для другой лошади "ли в других условиях. Только в строгом научном эксперименте могут быть вскрыты глубокие причины различных проявлений работоспособности рысистых лошадей, выработаны общие принципы и методы индивидуализации их тренировки.
В процессе исследований газоэнергетического обмена и дыхания у рысаков во время их движения, проведенных в лаборатории тренинга Института коневодства, было изучено возрастание энергетических затрат с повышением интенсивности тренировочных нагрузок. При этом выявлены закономерности дыхательной функции у рысаков, в том числе синхронность ритмов дыхания и движения. Нервнорефлекторные процессы синхронизации ритмов дыхания и движения были изучены с помощью специально сконструированной многоканальной аппаратуры. Широко исследовались также вопросы общей тренированности быстроаллюрных лошадей по гематологическим показателям и состоянию нервно-мышечного аппарата.
Физиологически обоснованный тренинг не может строиться на каких-либо отдельных, пусть даже важных, показателях состояния организма лошади. В современную физиологию прочно вошло понимание единства всех физиологических функций организма и его взаимосвязей с окружающей средой, единства, которое обеспечивается центральной нервной системой животного. С этих павловских позиций нервизма и следует рассматривать вопросы работоспособности рысистой лошади. Такой подход стал особенно необходимым после выявления в 1961 г. важнейшей физиологической закономерности высокой работоспособности рысака-синхронности ритмов дыхания и движения на резвом рысистом аллюре, в основе которой лежит координирующая роль центральной нервной системы. Следует отметить, что регуляторные функции центральной нервной системы рысистых лошадей изучены как в аспекте значимости ее основных типологических свойств, так и под углом зрения происходящих в ней функциональных изменений.
Широкие исследования высшей нервной деятельности рысаков позволили выявить зависимость характера их работоспособности от основных свойств центральной нервной системы, в частности возрастные особенности проявления резвости и предрасположенности к испытаниям на разные дистанции рысаков различного типа высшей нервной деятельности. Контроль за изменениями функционального состояния центральной нервной системы рысаков в процессе их тренинга, осуществляющийся по специально разработанной методике, привел к выяснению роли функциональной активности возбудительного и тормозного процессов для установления четкой синхронности ритмов дыхания и движения, устойчивости и резвости рысистого аллюра.
Данные разносторонних физиологических исследований в сочетании с анализом различных методов и практических приемов тренинга позволили уточнить основные физиологические принципы повышения резвости рысаков и выработать систему их тренировки. Эта система, предусматривающая индивидуализацию тренинга рысаков в соответствии с типологическими особенностями их высшей нервной деятельности, была детализирована и уточнена в процессе производственной апробации.
Чтобы правильно осмыслить рекомендуемые в настоящем пособии приемы тренировки, необходимо иметь хотя бы самое общее представление о закономерностях физиологических процессов в организме рысистой лошади в связи с ее тренировкой и испытаниями. Поэтому в первой главе книги в возможно более доступной форме изложены самые основные понятия о физиологии центральной нервной системы рысистых лошадей, особенностях их дыхания и движения и сущности энергетических процессов, протекающих в их организме. В этой же главе приводится и физиологическая характеристика всех элементов тренировки и испытаний рысаков.
Основываясь на глубоких теоретических положениях и накопленном за многие десятилетия опыте, в последующих главах пособия освещены вопросы заводского и ипподромного тренинга со всеми частными деталями (сборка, ковка и т. д.). В заключительных главах описываются деятельность ипподромов и зарубежный опыт испытаний рысаков. ___________________________________________________________________________
Глава 1. Физиологические основы тренировки рысистых лошадей
Основные понятия о физиологии центральной нервной системы рысистых лошадей
Организм лошади представляет собой сложную биологическую систему. Единство всех физиологических функций организма и его взаимосвязь с окружающей средой обеспечиваются центральной нервной системой, которая управляет деятельностью всех органов, посылая в них соответствующие импульсы возбуждения.
Физиология высшей нервной деятельности изучает закономерности работы центральной нервной системы, осуществляемой с помощью врожденных (безусловных) рефлексов и рефлексов, приобретенных в процессе индивидуального развития. Высшая нервная деятельность с ее временными условнорефлекторными связями - основа тончайшего приспособления организма лошади к меняющимся условиям внешней среды, основа всех приобретенных животным навыков, привычек, особенностей поведения и выездки. Постоянные, врожденные, связи между нейронами (нервными клетками) спинного мозга служат основой безусловных рефлексов; временные связи, образующиеся в коре больших полушарий головного мозга,- основа условнорефлекторных реакций.
Безусловные рефлексы не вырабатываются в процессе индивидуального развития организма. Они обусловлены видовыми биологическими особенностями животного и передаются по наследству. Путь безусловного рефлекса определяется строением нервных путей.
Например, болевое раздражение кожи животного воспринимается кожными рецепторами (чувствительными окончаниями нервных волокон); от них возбуждение по центростремительным нервам передается в центральную нервную систему, а затем на соответствующие центробежные нервные структуры и, приходя по ним к мышцам, приводит их в действие, в результате чего животное отстраняется от источника боли.
Условнорефлекторные связи возникают в процессе индивидуального развития животного в высших отделах его центральной нервной системы - в коре головного мозга.
Допустим, лошадь слышит скрип двери, открываемой в фуражную комнату. Слуховые рецепторы лошади воспринимают звуковое раздражение и передают возбуждение в соответствующий центр коры головного мозга - слуховой анализатор. Здесь это возбуждение сохраняется до того времени, пока наездник не принесет корм. Под действием пищи в организме возникает безусловный пищевой рефлекс: лошадь жует, выделяет слюну, глотает. В это время из нервных структур пищевого центра спинного мозга возбуждение поступает в соответствующий участок коры головного мозга - пищевой центр высшего коркового уровня. Таким образом, в коре головного мозга лошади возникают два очага возбуждения: один в слуховом центре от первого раздражителя (скрип двери); другой в пищевом центре в результате его возбуждения пищей. При слиянии этих двух возбуждений возникает временная условнорефлекторная связь.
После нескольких таких сочетаний скрип двери в фуражную становится для этой лошади сигналом пищевого условного рефлекса. Теперь уже само звуковое раздражение - скрип двери - вызывает у лошади пищевую реакцию.
Предположим далее, что наездник имеет обыкновение злоупотреблять хлыстом и держит его в фуражной. Тот же самый звуковой раздражитель - скрип двери - связывается уже не с пищевым, а с оборонительным центром, становится сигналом боли, и лошадь, услышав его, прядет ушами и уходит в дальний угол денника. Такой рефлекс потому и называется условным, что он образуется в определенных условиях и исчезает, если эти условия изменяются.
Для возникновения условного рефлекса требуется, чтобы ранее безразличный раздражитель несколько предшествовал по времени той деятельности, с которой он должен быть связан. Тогда этот раздражитель (например, скрип двери) становится сигналом определенной реакции и дает животному возможность подготовиться к соответствующей реакции.
Сигнальная роль тех или иных раздражителей имеет важнейшее биологическое значение.
Условные рефлексы, сигналами которых служат естественные свойства безусловных раздражителей, например вид корма, называются натуральными. По характеру безусловнорефлекторной основы, на которой выработан условный рефлекс, он обозначается как пищевой, оборонительный, ориентировочный и т. д. Кроме того, по названию анализатора, воспринимающего условный сигнал, условные рефлексы классифицируют на зрительные, слуховые, обонятельные.
Характер образования, функционирования и перестройки условнорефлекторных связей определяется свойствами возбудительного и тормозного процессов, взаимодействующих в центральной нервной системе лошади. Из исчисляемого миллиардами количества нервных клеток животного в каждый конкретный момент одни находятся в возбужденном состоянии, другие - в заторможенном, а какое-то количество клеток из возбужденного состояния переходит в тормозное, из тормозного - в возбужденное... Эту "мозаическую картину" И. П. Павлов называл "грандиозной динамической системой". Нервная система - это неоднородная, бесконечно сложная организация. Одни ее клетки, центры, отделы могут быть очень сильно возбуждены, в то время как другие заторможены.
Нервный центр - одно из основных функциональных понятий современной физиологии - представляет собой совокупность нервных клеток, деятельность которых необходима и достаточна для осуществления соответствующей функции.
Если лошадь бежит, то двигательный, дыхательный и ряд других центров возбуждены, а пищевой, половой и некоторые другие центры заторможены. Когда возбужден половой центр, то обычно бывает заторможен пищевой. Например, жеребец, находящийся в половом возбуждении, не будет есть поднесенный ему корм не потому, что не хочет, а потому, что не может, так как пищевой центр его в это время заторможен.
В центральной нервной системе животного и соответственно в ее высших отделах постоянно взаимодействуют возбудительные и тормозные процессы. Роль первых состоит в активации всякого рода деятельности. При этом нервная система управляет деятельностью всех эффекторов (каждым сокращением мышц, вдохом, глотанием, выделением каждой капли слюны и т. п.) посылом в них соответствующих импульсов возбуждения. Понимание же значения тормозных процессов намного сложнее. В коневодческой литературе роль их явно недооценивается и почти не освещена. И. П. Павлов указывал, что возбудительный и тормозной процессы одинаково важны, одинаково существенны в нервной деятельности.
Тормозной процесс выполняет три важнейшие роли - координационную, охранительную и восстановительную. Рассмотрим их на следующих примерах.
Предположим, лошадь сгибает ногу. Это движение сопряжено с сокращением мышц-сгибателей в результате возбуждения соответствующих нервных центров и расслаблением мышц-разгибателей вследствие торможения других центров, управляющих деятельностью этих мышц. Сгибание ноги сопряжено также с перемещением центра тяжести лошади, перераспределением нагрузок на другие ноги, большим напряжением одних мышц и расслаблением других, то есть со сложной координационной ролью центральной нервной системы. Акт движения связан не только с сокращением мышц, но и с их своевременным расслаблением.
Для осуществления дыхательной функции важно, чтобы возбуждению центра вдоха соответствовало торможение центра выдоха, а возбуждению центра выдоха - торможение центра вдоха.
Всякая полезная для организма деятельность может совершаться лишь тогда, когда вызывающие ее импульсы возбуждения целесообразно согласованы между собой. Эта координация возбуждений в центральной нервной системе и является главной функцией тормозного процесса. Чем интенсивнее совершается в организме какая-нибудь координированная деятельность (например, движение лошади резвой рысью), тем большего напряжения она требует не только от возбудительного, но и тормозного процесса.
Значение тормозного процесса как обязательного фактора координации впервые было показано И. М. Сеченовым. В случаях чередующегося ритма движений конечностей состоянию возбуждения центра одной из них сопутствует состояние торможения центра другой.
Охранительная, защитная роль торможения выражается в том, что торможению подвергаются различные функции организма, нецелесообразные в данной обстановке. Эта роль тормозного процесса наиболее ярко проявляется в условиях утомления, когда необходимо предохранить организм от перенапряжения.
К защитным функциям тормозного процесса примыкают его восстановительные функции. Ученик И. П. Павлова академик Г. В. Фольборт пришел к выводу о том, что повышение работоспособности при чередующейся работе конечностей является следствием повторяющихся кратковременных актов торможения в работающих центрах и что торможение является стимулятором восстановительных процессов.
Координационные, защитные и восстановительные функции тормозного процесса теснейшим образом связаны между собой. Эти функции тормозного процесса в современной физиологии стали вопросами широкого и конкретного практического действия.
Таким образом, сокращение какой-либо мышцы и вообще деятельность любого органа обусловливаются возбужденным состоянием соответствующего центра, посылающего к мышце (органу) импульсы возбуждения. Когда же этот центр, то есть группа клеток в центральной нервной системе, переходит в заторможенное состояние, то соответствующая мышца расслабляется.
Представим себе лошадь, бегущую резвой рысью. Сотни ее мышц и сухожилий действуют координирование, то есть не сами по себе, а согласованно. При этом одни мышцы сокращаются, другие расслабляются; в следующий момент действует уже иная комбинация. Механическая борьба мышц на периферии отсутствует, она заменена внутрицентральной борьбой в виде столкновения процессов возбуждения и торможения.
Итак, различные участки центральной нервной системы животного взаимодействуют между собой. Основные способы такого взаимодействия заключаются в следующем. Когда какой-либо центр возбужден, то возбуждение распространяется сначала на ближайшие к нему участки мозга, а затем и на более дальние. Этот процесс называется иррадиацией возбуждения. Торможение тоже имеет свойство иррадиировать.
Пример наиболее иррадиированного возбуждения - предстартовое состояние лошади, а иррадиированного торможения (разлитое торможение) - сон.
Иррадиации противоположно явление концентрации нервного процесса, то есть усиление его активности на ограниченном участке нервной ткани. Возбуждение ограничивается торможением, торможение - возбуждением. Концентрация возбуждения или торможения - результат взаимодействия обоих этих процессов. При напряженной координированной деятельности, например на резвой рыси, возбудительные и тормозные процессы взаимодействуют в резко концентрированной форме. Концентрация процессов, особенно быстрая их концентрация,- дело трудное, требующее от центральной нервной системы большого напряжения и тренированности. При этом особую трудность для нервной системы представляет быстрая концентрация тормозного процесса. Этим и объясняется сложность управления лошадьми на старте, то есть в момент, когда возбуждение под влиянием массы внешних раздражителей широко иррадиирует по коре головного мозга лошади, а торможение не успевает достаточно сконцентрироваться, чтобы целесообразно его ограничивать.
Фактически каждое проявление какой-либо реакции, каждый акт жизнедеятельности лошади является результатом взаимодействия возбудительного и тормозного процессов в ее центральной нервной системе. В результате взаимодействия нервных процессов осуществляется и образование условных рефлексов.
До сих пор речь шла об активном тормозном процессе, протекающем в коре головного мозга животного, так называемом внутреннем торможении. Важнейшая черта его - закономерность возникновения внутри центральных нервных структур для обеспечения целесообразной жизнедеятельности организма.
В деятельности центральной нервной системы проявляется и так называемое внешнее торможение.
Представим себе довольно типичную картину. В новой обстановке лошадь словно оцепенела; заданный ей корм, который она любит, не ест, словно не видит его. Внешний фактор (новая обстановка) так подействовал на нервную систему лошади, что затормозились полезные для организма функции (лошадь не может есть), затормозилась и сама возможность нервной системы гибко управлять действиями организма.
Характерная черта внешнего торможения состоит в его неактивности, это как бы подчинение нервной системы животного действию внешнего фактора.
Наиболее опасно так называемое запредельное внешнее торможение, возникающее под действием сверхсильного (для данной лошади) раздражителя. Оно может иметь самые неожиданные последствия.
Например, лошадь испугалась проехавшего вблизи нее бульдозера и "подхватила на унос". Почему она не подчиняется действию обычных, хорошо ей знакомых средств управления? Потому что условные рефлексы, на которых базировались ее выездка - подчинение и послушание, отключены; они подверглись действию внешнего торможения.
Лошадь вся "в мыле", но продолжает бешеную скачку. С предельным напряжением работают все ее органы, кроме одного - коры головного мозга. В низших отделах ее центральной нервной системы, особенно в центрах, связанных с пассивно-оборонительной реакцией, небывалое, сверхсильное возбуждение. Случай, когда низшие отделы центральной нервной системы выходят из-под контроля коры головного мозга, И. П. Павлов называл "бунтом подкорки".
Допустим, на пути такой лошади стоит забор. Когда в спокойной обстановке перед лошадью возникает непреодолимая преграда, у нее включаются натуральные защитные условные рефлексы, срабатывает внутренее торможение и она останавливается или сворачивает в сторону. Запредельное внешнее торможение отключает всякие корковые регуляции и в первую очередь те, которые связаны с активностью внутреннего торможения. Поэтому в приведенной здесь ситуации лошадь может даже налететь на забор. Внешнее торможение явится в этом случае причиной самого нецелесообразного действия животного.
От исследователя, тренера и вообще специалиста-коневода требуется четко разграниченное понимание сущности внутреннего и внешнего торможения. Когда речь идет о внутреннем торможении, его можно назвать активным, можно корковым, а можно и просто тормозным процессом. Когда же речь заходит о внешнем торможении, а тем более о запредельном внешнем торможении, то термины "внешнее" и "запредельное" обязательны. Из приведенного выше примера следует, что запредельное внешнее торможение не во всех случаях связано с прекращением движения лошади. Для него характерно прекращение нормального функционирования высших отделов ее центральной нервной системы: кратковременное или длительное, частичное или более полное их отключение от руководства организмом, что выражается в выпадении из поведения лошади навыков, выработанных на условнорефлекторной основе.
По комплексу основных свойств нервной системы - силе нервных процессов (возбудительного и тормозного), их уравновешенности и подвижности - лошади могут быть отнесены к тому или иному типу высшей нервной деятельности. Сила нервной системы выражается в ее способности выносить значительное напряжение возбудительного и тормозного процессов, сохраняя при этом нормальную работоспособность. Из постоянного и правильного балансирования возбудительного и тормозного процессов складывается нормальная жизнь животного.
Подвижность нервных процессов определяет быстроту образования и перестройки условнорефлекторных связей. Исследования показали, что среди лошадей, как и среди животных других видов, можно выделить представителей четырех классических павловских типов высшей нервной деятельности (ВНД*) (табл. 1).
Нервная деятельность у лошадей сильного уравновешенного подвижного типа отличается активностью и устойчивостью. Условнорефлекторные связи быстро возникают и при необходимости быстро перестраиваются.
У лошадей сильного уравновешенного инертного типа нервная система сильная и хорошо уравновешенная, однако условнорефлекторные связи возникают и перестраиваются медленнее. Инертность нервных процессов в физиологическом смысле означает медленное привыкание к чему-либо, медленное освоение с новой обстановкой, то есть медленное образование условных рефлексов. Поэтому нет ничего странного в том, что среди резвейших быстроаллюрных лошадей есть животные сильного уравновешенного инертного типа.
Неуравновешенность нервной системы выражается в преобладании по силе возбудительного процесса над тормозным. У некоторых лошадей над сильным тормозным процессом преобладает сильнейший возбудительный. Таких животных И. П. Павлов называл слишком сильными. У других неуравновешенность является следствием недостаточной силы тормозного процесса.
У лошадей с неуравновешенной нервной системой затруднены те формы нервной деятельности, которые требуют концентрации, быстрой мобилизации или большого напряжения тормозного процесса.
Лошадям слабого типа ВНД свойственна слабость и возбудительного, и тормозного процессов. Слабость нервной системы лошади выражается в том, что она не в состоянии переносить значительное напряжение возбудительного и тормозного процессов; в результате перенапряжения нервная система таких животных подвергается действию внешнего торможения. ___________________________________________________________________________
Особенности взаимодействия возбудительного и тормозного процессов в центральной нервной системе рысистой лошади
Движение лошади резвой рысью вызывает значительное напряжение возбудительного и тормозного процессов ее центральной нервной системы, причем на разных этапах дистанции условия взаимодействия этих процессов меняются, что необходимо учитывать и при езде на приз, и в процессе тренировки рысаков. Особенностями взаимодействия нервных процессов во многом и определяются дистанционные способности рысистой лошади.
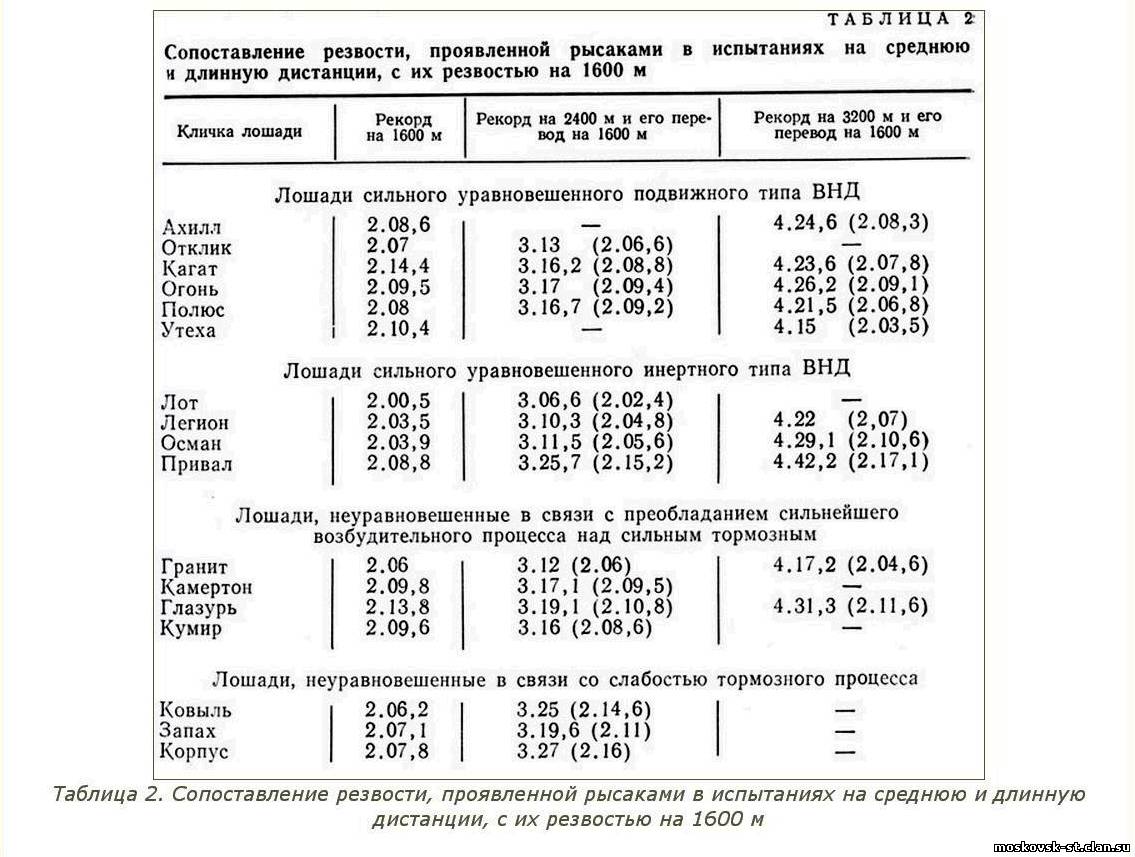
На развитие дистанционных способностей лошадей, кроме системы тренировки, влияет и врожденная ее предрасположенность. Одни лошади проявляют максимальную резвость в испытаниях на короткие дистанции, другие - на длинные. Склонность к проявлению максимальных резвостных возможностей на определенных дистанциях, - как правило, достаточно стабильное в течение всей беговой карьеры качество лошади. Показателем дистанционных способностей служит не абсолютная резвость лошади, зависящая от ее резвостного класса, а сравнение резвости, показанной ею на разных дистанциях. Такое сравнение возможно путем пересчета резвости, проявленной рысаком на средней и длинной дистанциях, в эквивалентную резвость на 1600 м. Для этого пользуются специальными таблицами, составленными с учетом утомляемости лошади.
Настоящими дистанционерами следует считать рысаков, способных бежать основные дистанции (1600, 2400, 3200 м) в одну и ту же резвость (с учетом поправки на утомляемость). У лошадей с плохими дистанционными способностями резвость обычно не удерживается на одном уровне даже на дистанции 1600 м, и последнюю четверть круга они пробегают, как правило, медленнее. К лошадям средних дистанционных способностей можно относить таких, которые хорошо бегут на 1600 и 2400 м, могут выдерживать и более длинные дистанции, но с меньшей результативностью, чем истинные дистанционеры. Предрасположенность рысистых лошадей к проявлению высокой резвости в испытаниях на определенные дистанции более рельефно выступает у наиболее классных лошадей старшего возраста.
На короткие дистанции, вопреки мнению, распространенному в коневодческой литературе, наиболее успешно бегут рысаки, нервной системе которых свойственны сила и уравновешенность возбудительного и тормозного процессов при их инертности. В беге на длинные дистанции хорошо выступают рысаки сильного неуравновешенного типа высшей нервной деятельности, у которых над сильным тормозным процессом преобладает сильнейший возбудительный.
Как понять взаимосвязь этих явлений? На старте нервная система лошади подвергается множеству внешних возбуждающих влияний: различные условнорефлекторные сигналы предстартового состояния, пробегающие мимо лошади, обстановка беговой дорожки, музыка и шум на трибунах, заполненных публикой, и т. д. Возбудительный процесс у лошадей быстро развивается и распространяется (иррадиация), а обстановка на старте содействует этому. Тормозной же процесс у лошадей протекает значительно медленнее и в условиях старта не стимулируется окружающей обстановкой. Таким образом, слаженность взаимодействия процессов в центральной нервной системе рысистой лошади на старте затрудняется в результате ее общего перевозбуждения, что легко может привести к нарушению координации движений - снижению резвости, сбою или проскачке. А значение хорошо принятого старта наиболее велико при испытаниях на короткую дистанцию.
Становится понятным, почему наибольший успех на коротких дистанциях сопутствует лошадям, нервная система которых характеризуется замедленным развитием возбудительного процесса, то есть лошадям сильного уравновешенного инертного типа ВНД. Их нервная система обладает наилучшими возможностями для установления сбалансированного взаимодействия обоих процессов и принятия резвого старта.
Другая ситуация создается к концу длинных дистанций, когда под влиянием сильного общего утомления в центральной нервной системе рысистой лошади начинает преобладать охранительная роль широко иррадиированного тормозного процесса, препятствующего поддержанию высокой резвости. На финише длинных дистанций повышенная сила возбудительного процесса позволяет преодолевать утомление и способствует мобилизации всех сил и возможностей организма. Этим объясняются успешные выступления в беге на длинные дистанции лошадей сильного неуравновешенного типа высшей нервной деятельности, у которых над сильным тормозным процессом преобладает сильнейший возбудительный.
С рассмотренных позиций становится совершенно понятным, почему лошади с сильными уравновешенными подвижными нервными процессами успешно выступают на всех дистанциях, а лошади, у которых слаб тормозной процесс или слабы оба процесса, нестабильны по результативности и почти не выступают на длинных дистанциях (табл. 2).
Таким образом, предрасположенность рысистых лошадей к развитию дистанционных способностей определяется типом их высшей нервной деятельности.
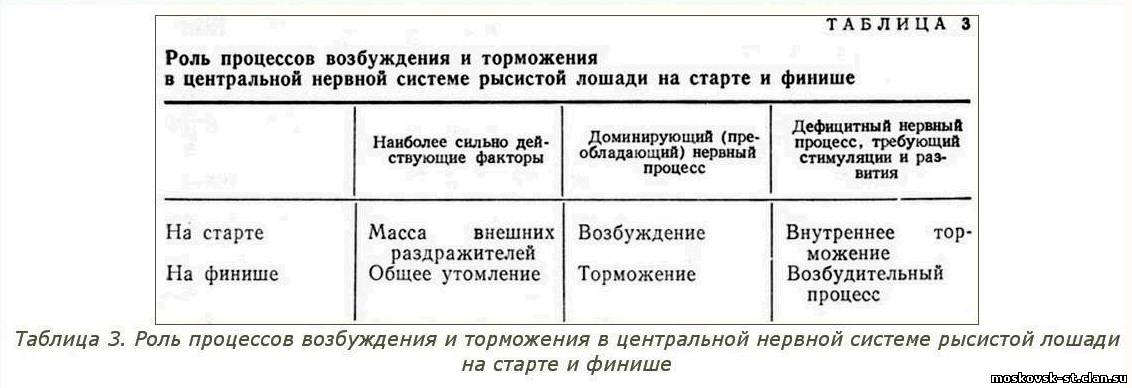
В главе "Индивидуализация тренировки и испытаний рысистых лошадей в соответствии с типом их высшей нервной деятельности" будут подробно рассмотрены приемы отработки четкого старта и резвого финиша у лошадей разного типа ВНД. Здесь же в таблице 3 приведены лишь основные закономерности.
На старте нервную систему рысистой лошади возбуждать не следует, поскольку под влиянием предстартовой обстановки она и так возбуждена. Поэтому не рекомендуется применять ни резкий посыл, ни сдерживание лошади жестким управлением. При туго натянутых вожжах удила причиняют лошади боль и возбуждают ее нервную систему. При отработке резвого приема на тренировках необходимо мягкое спокойное управление лошадью.
Резвость бега в конце дистанции поддерживается усилением возбудительного процесса, который следует всячески стимулировать. В частности, при езде на приз прибегают к энергичному посылу лошади на финише с применением в случае необходимости даже болевых раздражений хлыстом. Во время тренировки важно вырабатывать у лошади устойчивый рефлекс броска на финише. Для этого на маховых и резвых работах последняя четверть должна быть всегда самой резвой. В результате обычные на финишной прямой раздражители приобретают значение условных сигналов для повышения активности возбудительного процесса и проявления резвого броска на финише, столь необходимого для одержания победы над соперниками. ___________________________________________________________________________
Сущность энергетических процессов в организме лошади при ее движении
Энергия абсолютно необходима для любых проявлений жизнедеятельности, тем более необходима она для всякого движения живых существ.
Движение лошади резвой рысью сопряжено со значительным повышением интенсивности энергетических процессов в ее организме. Во время движения в результате трансформации энергии, аккумулированной в организме, лошадь совершает механическую работу по преодолению силы тяжести, инерции, силы трения, сопротивления воздуха и т. п. Энергия же существует в двух видах: как энергия движения, или кинетическая, и как энергия покоя, или потенциальная. При движении тела какое-то количество потенциальной энергии переходит в Кинетическую.
Внешние силы сопротивления движению лошадь преодолевает соответствующей механической работой. Эта работа осуществляется в результате расхода внутренней энергии, аккумулированной в тканях организма лошади в виде сложных химических соединений.
К мышцам, выполняющим при сокращении внешнюю механическую работу, притекает богатая кислородом артериальная кровь; от мышц же оттекает по венам кровь, обедненная кислородом и содержащая продукты распада, главным образом углекислый газ, выделяющийся затем с выдыхаемым лошадью воздухом. Таким образом, напряженная мышечная работа при движении лошади резвой рысью требует интенсивного поступления через легкие в кровь кислорода и его доставки к работающим мышцам, то есть требует соответствующей тренированности дыхательной и сердечно-сосудистой систем. Взаимодействие физиологических систем, участвующих в энергетически мощной деятельности организма, регулируется центральной нервной системой лошади и гуморально, то есть соответствующими изменениями состава крови.
Начало движения сопряжено с возбуждением центральной нервной системы лошади. Возбуждается двигательный центр, затем сразу же дыхательный центр, стимулирующий дыхание. Возбуждаются центры вегетативной нервной системы, управляющей деятельностью внутренних органов и стимулирующей работу сердца. Стремительно увеличиваются потребление лошадью кислорода из воздуха и его доставка к работающим мышцам. Но мышцы расходуют такое количество кислорода, что кровь все же обедняется им, в результате чего в ней накапливается много углекислого газа, являющегося сильнейшим гуморальным раздражителем дыхательного центра. Омывая дыхательный центр, такая кровь стимулирует дыхание, и оно становится более продуктивным. Тем не менее, если бы кровь не обладала буферными свойствами, концентрация углекислого газа в ней быстро стала бы чрезмерной.
Буферные системы крови - это комплексы растворенных в ней химических соединений, способных связать и на какое-то время нейтрализовать вредные для организма вещества. Развитие буферных систем крови имеет особенно большое значение для периодов резкого возрастания энергетических процессов и преобладания их над дыхательной функцией. Применяемые в системе тренинга рысистой лошади короткие резвые броски способствуют развитию буферных систем крови.
Структуры буферных систем нормализуются в процессе выделения из них углекислоты. Поэтому после прекращения интенсивного движения дыхательная и кровеносная системы лошади еще продолжают некоторое время напряженно функционировать, лошадь продолжает потреблять из воздуха большое количество кислорода и выделять много углекислого газа. Это длится до тех пор, пока не выделится связанная буферными системами углекислота и не восполнятся кислородные ресурсы крови и мышечных тканей, то есть пока в организме лошади не будет ликвидирована временная кислородная задолженность.
Величина кислородной задолженности (кислородного долга) определяется количеством кислорода, израсходованным при движении лошади из внутренних резервов ее организма и восполняющимся из атмосферного воздуха после прекращения интенсивного движения.
При длительном интенсивном движении резвой рысью мобилизуются все ресурсы организма: на полную мощность работают дыхательная и кровеносная системы, расходуются резервы гликогенных депо и максимально используются возможности буферных систем крови. Таким образом, при движении резвой рысью энергетически мощные процессы, протекающие преимущественно в мышцах лошади, обеспечиваются деятельностью всего ее организма. Следовательно, по показателям уровня энергетических процессов в организме лошади при ее движении тем или иным аллюром можно судить не только о работе мышц, но и о напряженности различных физиологических процессов во всем ее организме.
Физиология рысистой лошади на современной научной основе стала доступна пониманию лишь после того, как была изучена динамика энергетических процессов в организме рысаков во время их движения с разной скоростью. Это было сделано на Московском ипподроме в 1960 - 1961 гг. с помощью специально разработанной методики и аппаратуры (рис. 1).
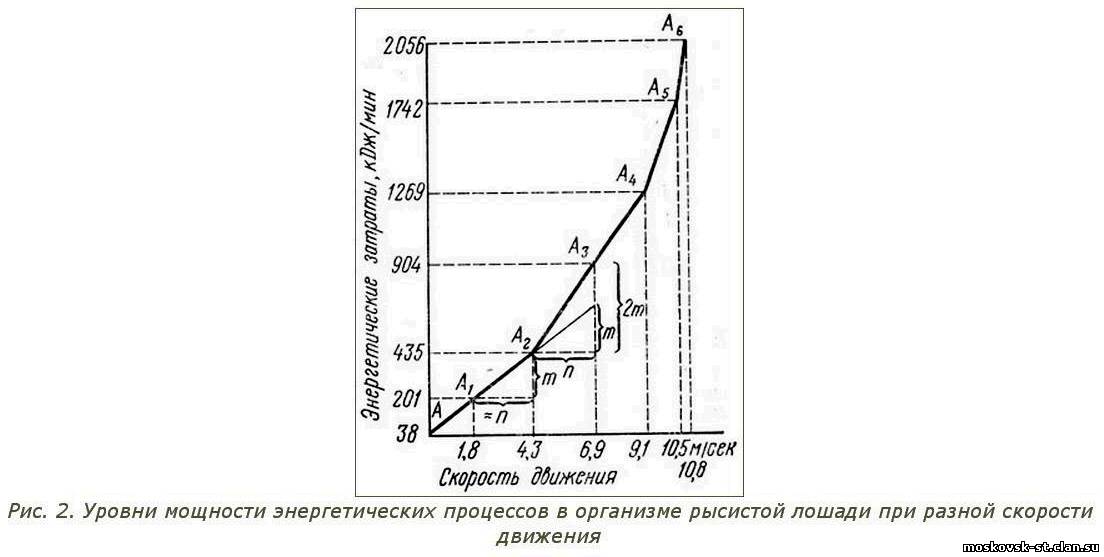
Нарастание энергетических процессов в организме рысистых лошадей при разной скорости их движения графически изображено на рисунке. 2. Из этого графика следует, что при движении шагом энергетические процессы в организме лошади протекают примерно в 5 раз, а при движении тротом - в 11 раз интенсивнее, чем в состоянии относительного покоя, а темпы прироста скорости движения приблизительно пропорциональны темпам прироста мощности энергетических процессов. Это свидетельствует о том, что движение рысистой лошади шагам и тротом совершается при одинаковом коэффициенте полезного действия энергетических затрат.
Рассмотренная нами пропорциональность отражена на графике прямизной линии АА2.
Неизменность коэффициента полезного действия энергетических затрат свидетельствует о том, что движение шагом и тротом совершается при оптимальном режиме работы организма. Оба эти аллюра естественные, врожденные и привычные для лошади; они не сопровождаются образованием в ее организме кислородной задолженности и характеризуются рядом других существенных особенностей. Благодаря этому тихие работы являются важнейшей составной частью системы тренировки рысаков.
При движении рысака размашкой коэффициент полезного действия приращенных энергетических затрат снижается почти вдвое в точке А2 кривая пошла вверх и почти не (меняет угла наклона вплоть до точки А4. Приращение скорости движения на размашке относительно скорости движения тротом равно 2,6 м/с (6,9 - 4,3 м/с) и на графике соответствует величине n. Если бы коэффициент дополнительных энергетических затрат оставался прежним, то их прирост при переходе с трота на размашку был бы равен величине т. Фактически же увеличение дополнительных энергетических затрат вдвое превышает величину га и выражается на графике вели8чиной 2 m.
Таким образом, увеличение скорости движения (допустим, на 1 м/с; на размашке и на махе достигается вдвое большими дополнительными затратами энергии, чем на троте. Увеличение энергетических затрат для приращения скорости лишь в незначительной мере может быть объяснено возрастанием сопротивления воздуха и других механических факторов. Существо же явления заключается в том, что при высокой интенсивности энергетических процессов (на размашке в 24 раза, а на махе в 33 раза интенсивнее, чем в покое) изменяется характер биохимических реакций, резко возрастает теплопродукция, увеличиваются потери тепла и организм лошади функционирует уже в менее экономном режиме, чем на шагу и троте. На размашке и махе организм рысака находится в более трудных условиях и вынужден переносить их, не снижая скорости движения.
Примечательно, что при увеличении интенсивности энергетических прецессов на махе в 1,5 раза по сравнению с размашкой темпы прироста энергетических затрат, необходимых для приращения скорости движения, остаются такими же (прямая линия А2А4). Следовательно, при движении лошади махом физиологические процессы протекают интенсивнее, чем при размашке, но без резкого качественного скачка ,(в точке А3), подобного скачку при переходе от трота к размашке (в точке А2). Движение резвой рысью тоже можно рассматривать как тренировочное, но это будет уже тренировка организма в еще более напряженном режиме. Прирост скорости при переходе от маха к резвой сопровождается увеличением в 2 раза энергетических затрат (на каждую долю секунды приращения скорости), а при переходе от резвой к более резвой - уже в 3 раза. Кривая графика сначала круто (А4А5), а затем почти отвесно (А5А6) поднимается вверх.
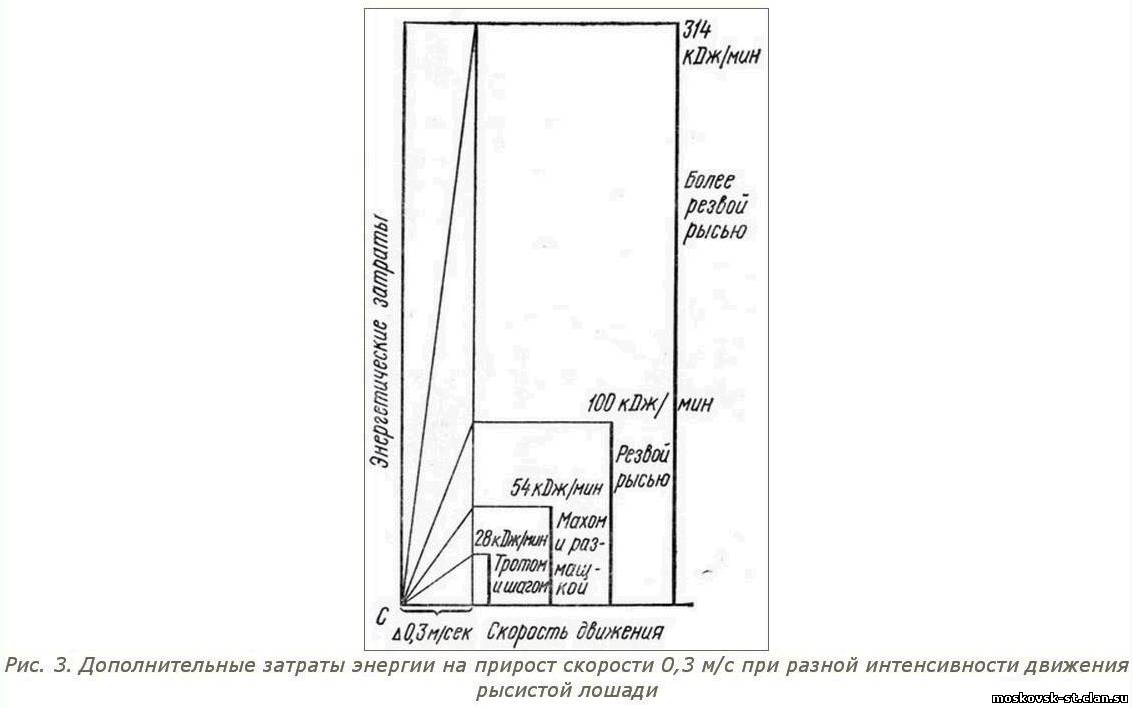
При движении резвой рысью приращение скорости (допустим, 0,3 м/с) сопровождается в 3 раза большими затратами энергии, чем при более тихой резвой рыси, в 6 раз большими, чем при движении махом, и в 12 раз большими, чем при движении тротом.
Графически это представлено на рисунке 3, причем соотношение скорости движения (по горизонтальной оси) и энергетических затрат (по вертикальной оси) остается тем же, что и на рисунке 2.
В точке С графика исходные показатели скорости движения лошади совмещены с энергетическими затратами в ее организме при движении шагом, тротом, размашкой и т. д. По горизонтальной оси отложено приращение скорости движения на 0,3 м/с. Из точки С лучеобразно поднимаются (под теми же углами, что и на рисунке 2) линии, характеризующие темпы повышения мощности энергетических процессов при увеличении скорости движения лошади.
Таким образом, если лошадь движется шагом или тротом, то при повышении скорости ее движения на 0,3 м/с мощность энергетических процессов в ее организме увеличивается на 28 кДж/мин. При движении лошади размашкой или махом этот показатель равен 54 кДж/мин, при движении резвой рысью - 100 кДж/мин, а при более резвой рыси - 314 кДж/мин. Безусловно, с несравненно еще более высоким напряжением энергетических и вообще всех физиологических процессов вынужден функционировать организм рысистой лошади при достижении предельной для нее скорости движения.
Организму всех животных свойственна тенденция расходовать энергию с возможно более высокой степенью эффективности. Физиологические процессы, протекающие в организме рысистой лошади, направлены на формирование такого режима работы, при котором максимальная скорость бега достигалась бы при минимально возможных энергетических затратах. Биологическая целесообразность этого заключается главным образом не в экономии энергии, а в предотвращении предельной, близкой к катастрофической перегрузки в первую очередь дыхательной, сердечно-сосудистой, кровеносной, выделительной и терморегуляторной систем рысистой лошади, обеспечивающих интенсификацию энергетических процессов.
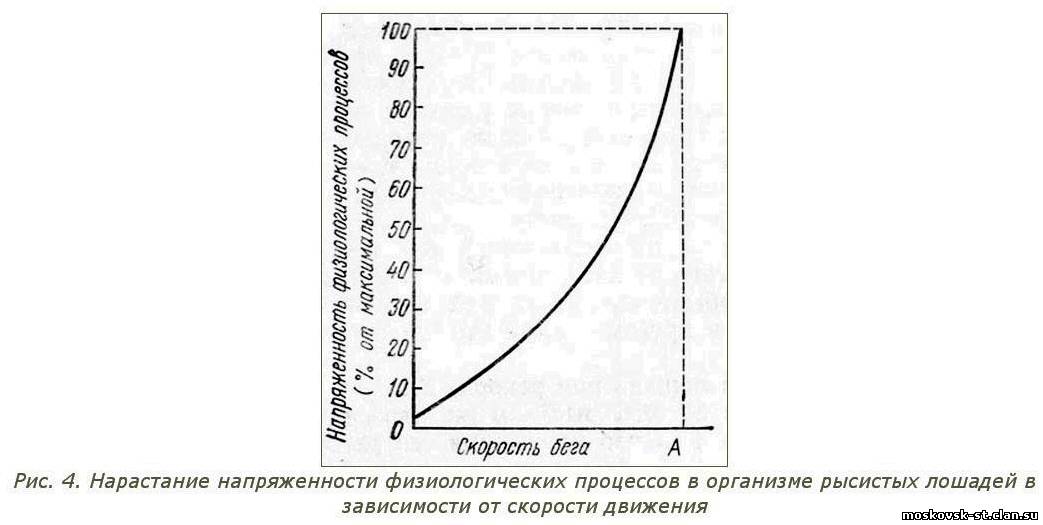
При общей тенденции организма рысистой лошади к минимальному возрастанию энергетических затрат они в условиях форсированного режима на резвой рыси все же чрезвычайно резко повышаются, создавая физиологический барьер дальнейшему наращиванию скорости движения. Уровень энергетических процессов отражает напряженность важнейших физиологических функций организма рысистой лошади, и его непомерное возрастание служит основой естественного, физиологического ограничения резвости ее бега, а следовательно, и ограничения прогресса резвости породы. О стремительном нарастании напряженности энергетических процессов с повышением скорости движения следует помнить при рассмотрении повседневных вопросов тренинга рысистых лошадей.
У какой лошади более выражен прогресс резвости? У той, которая пробежала 1600-метровую дистанцию за 2 мин 07 с (2.07) вместо 2 мин 15 с (2.15), или у пробежавшей ее за 2.05 вместо 2.10? В одном случае сброшены 8 с, в другом - 5, но это произошло при разных уровнях резвости.
Нередки случаи, когда двухлетняя лошадь при рекорде 3.15 на 1600 м, в течение месяца сбрасывает 30 с. С рысаком же класса 2.05 иногда работают в течение года с целью улучшения его рекорда на 1 с.
При рекорде трехлетней рысистой лошади, равном 2.20, резвую работу проводят примерно в 2.30, а маховую - в 2.45; при рекорде же, равном 2.10, резвую - в 2.17, маховую - в 2.35.
Следовательно, вопросы, касающиеся прогресса резвости рысаков и нормирования интенсивности их тренировочных работ, надо решать в зависимости от уровня, на котором происходит прогресс резвости, или от резвостных возможностей рысака в данное время, выражающихся его рекордом.
Рассмотренный выше график (рис. 2) позволяет сопоставлять интенсивность тренировочных нагрузок с учетом соответствующих уровней резвости. Но этот график справедлив лишь для лошадей класса 2.20 - 2.15, на базе исследования тренировочных нагрузок которых он построен. Для лошадей другого резвостного класса показатели энергетических затрат будут другими, но физиологическая закономерность их с приростом или уменьшением скорости движения останется аналогичной. Следовательно, она может быть выражена подобным же графиком, лишь сдвинутым несколько вправо или влево. Графическая, кривая, освобожденная от конкретных значений скоростей движения и воспроизведенная с овальными переходами в узловых точках, представляет собой достаточно точную графическую модель нарастания напряженности физиологических процессов у рысистых лошадей любого возраста и резвостного класса (рис. 4).
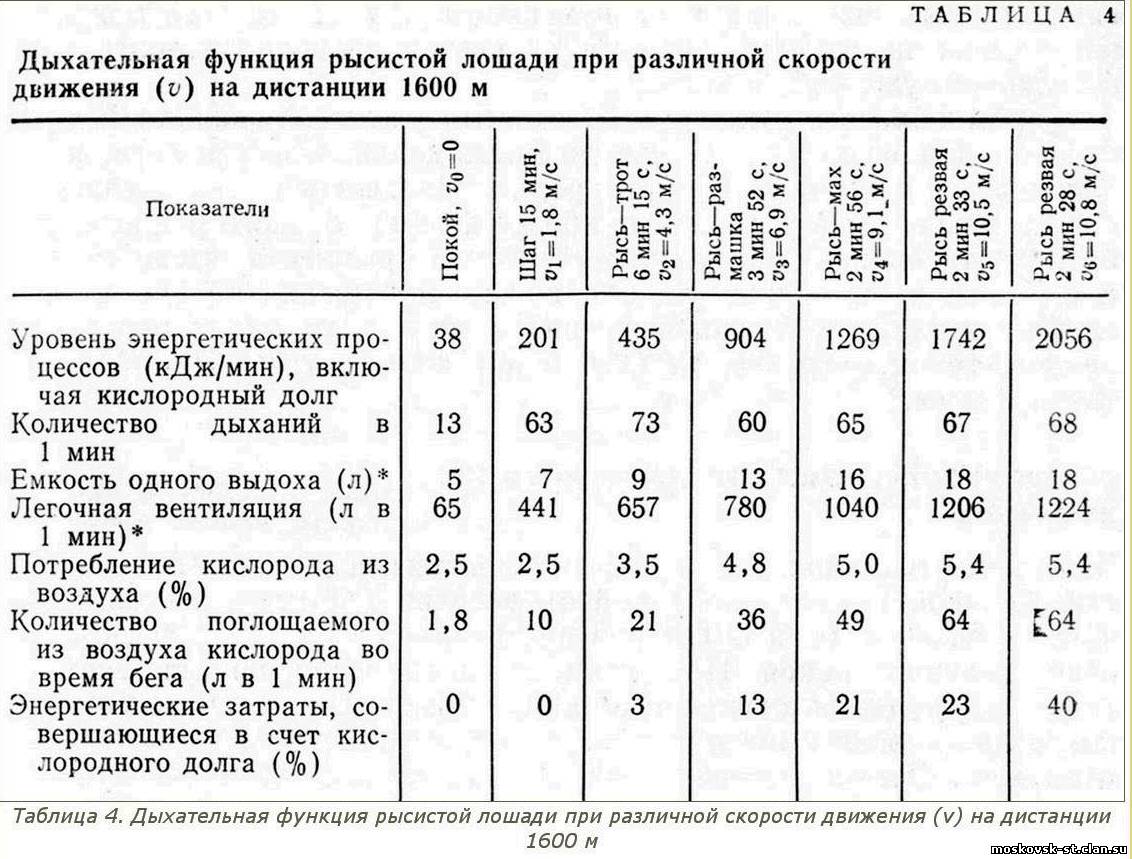
Что касается дыхательной функции рысистой лошади при разных уровнях энергетических процессов, связанных с разной скоростью движения, то соответствующие данные об этом представлены в таблице 4.
* (Показатели даны в пересчете на сухой воздух при 0°С и давлении, равном 760 мм ртутного столба. Фактически легочная вентиляция примерно на 10 - 20% больше.)
Легочная вентиляция, то есть количество воздуха, проходящего через легкие в 1 мин, зависит от частоты дыхания и емкости каждого вдоха (выдоха). По сравнению с покоем дыхание лошади на шагу резко учащается, а затем до движения резвой рысью хотя и колеблется по частоте, но возрастает не очень резко. Другое дело емкость одного выдоха, которая, как это следует из данных таблицы 4, при увеличении скорости движения лошади неуклонно возрастает. Следовательно, учащение дыхания представляет собой первый быстро включаемый источник увеличения легочной вентиляции, который исчерпывает свои возможности уже при движении лошади тротом. Увеличение же емкости одного выдоха - физиологически более глубокий источник, мощность которого наращивается постепенно от покоя к шагу, троту и далее до резвой рыси и за счет которого неуклонно увеличивается легочная вентиляция, достигая на резвой рыси 1200 л в 1 мин и приближаясь у некоторых лошадей к 2000 л в 1 мин.
Организм любой лошади, в первую очередь самой нетренированной, отвечает на переход к движению учащением дыхания. Более глубокое дыхание свойственно лишь хорошо тренированным лошадям. Поэтому одна из главных целей тренировки рысистых лошадей- увеличение легочной вентиляции в результате перехода на открытое глубокое, емкое дыхание. Интенсивность дыхания лошади увеличивается при усилении легочной вентиляции и повышении доли кислорода, поглощаемого легкими из каждого литра проходящего через них воздуха. При этом второй источник обеспечения кислородом энергетических процессов наиболее глубокий, но, как следует из данных таблицы 4, он начинает использоваться организмом не сразу. Значение его возрастает по мере повышения энергетических процессов в организме лошади: уже при движении ее махом из каждых 100 л воздуха, проходящих через легкие, поглощается вдвое больше кислорода, чем в состоянии покоя и при движении шагом, а на резвой рыси его поглощается еще больше.
Итоговым показателем интенсивности дыхания, отражающим эффективность физиологических процессов, направленных на возможно более полное снабжение кислородом организма движущейся лошади, служит количество кислорода, поглощаемого легкими из воздуха. По сравнению с состоянием покоя при движении шагом в легких лошади поглощается из воздуха в 5,5 раза, а при движении тротом - в 12 раз больше кислорода, то есть показатель этот увеличивается пропорционально энергетическим затратам.
При этом потребность организма в кислороде за счет дыхания полностью удовлетворяется и кислородной задолженности не наблюдается.
При движении размашкой энергетические затраты организма увеличиваются вдвое по сравнению с затратами энергии при троте (904 кДж по сравнению с 435 кДж), а кислорода в легких поглощается лишь в полтора раза больше (36 л по сравнению с 21 л). И несмотря на то что по сравнению с состоянием покоя поглощение 'кислорода увеличилось в 20 раз (36 л по сравнению с 1,8 л), снабжение организма движущейся лошади кислородом начинает отставать от уровня энергетических затрат. В результате этого в организме рысака нарастает кислородный долг, а потому включаются буферные и резервные системы организма. При движении лошади махом эти явления выражены еще резче и кислородный долг уже достигает 21%.
При движении резвой рысью в легких лошади поглощается из воздуха в 36 раз больше кислорода (до 64 л), чем в состоянии покоя, но энергетические затраты при этом возрастают в 46 раз (до 1742 кДж), так что кислородный долг продолжает увеличиваться.
Когда же рысь становится еще более резвой, то это сравнительно небольшое увеличение скорости достигается возрастанием в 54 раза энергетических затрат (до 2056 кДж). А так как интенсивность дыхания лошади не изменяется (64 л), то весь прирост энергетических затрат относится на счет кислородного долга, достигающего уже 40% и более.
Таким образом, дыхательная система рысистой лошади исчерпывает свои возможности еще до достижения резвости бега, предельной на данном этапе тренировки. Предельная же резвость бега развивается в условиях наиболее форсированного режима работы важнейших физиологических систем организма рысистой лошади. Это не причинит ущерба организму рысака только в том случае, когда все физиологические системы его будут подготовлены к такой нагрузке, то есть будут соответствующим образом тренированы. ___________________________________________________________________________
Синхронность ритмов дыхания и движения у рысаков
В периоды наивысшего напряжения организма рысака (при резвости, близкой к рекордной) расход энергии достигает максимального уровня - в 60 и более раз превышает ее расход в состоянии относительного покоя. При столь высокой интенсивности энергетических процессов организм лошади испытывает особенно острую потребность в кислороде и достаточно эффективном выделении продуктов обмена, в первую очередь углекислого газа. Дыхание в этих условиях приобретает совершенно исключительное значение. При отставании дыхательной функции от возрастающей потребности организма в кислороде образуется нарастающий кислородный долг, являющийся одним из решающих факторов, ограничивающих возможность интенсивного движения.
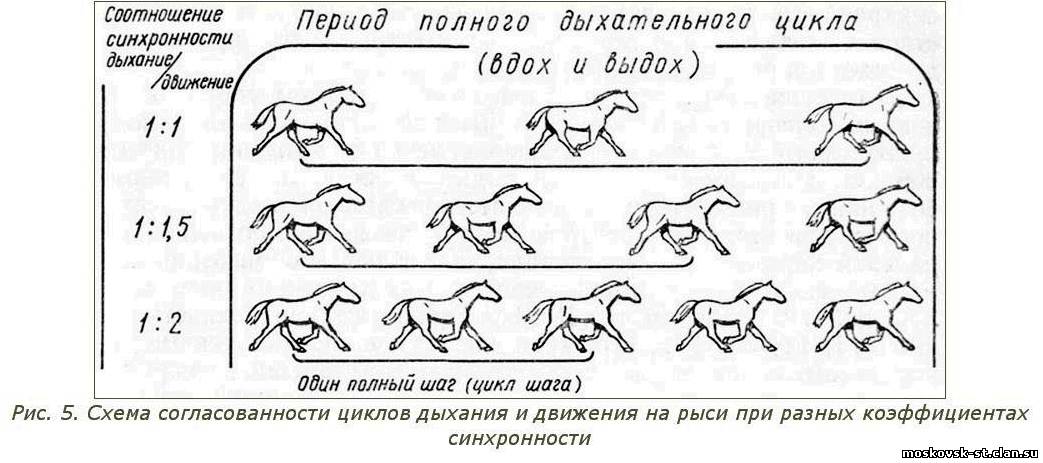
Как уже отмечалось, чем глубже дыхание, тем больше воздуха проходит через легкие и тем больше кислорода усваивается из каждых 100 л вдыхаемого животным воздуха. В результате этого эффективность снабжения организма лошади кислородом повышается. Правильное глубокое дыхание возможно лишь при синхронности ритмов дыхания и движения, то есть согласованности во времени циклов дыхания и движения (рис. 5). При синхронности этих циклов движения корпуса и конечностей облегчают дыхательные движения грудной клетки, а при разлаженности циклов затрудняют их. На рысистом аллюре соотношение частоты дыхания и частоты шагов рысака может быть разным. На тихой рыси оно обычно равно 1:1, то есть количество дыханий в минуту (например, 88) соответствует такому же количеству шагов. При. резвой рыси частота шагов увеличивается, а дыхание может становиться более редким и глубоким. Соотношение частоты дыхания и частоты шагов может составлять 1:1,5 или 1:2.
Если рысак делает вдох, предположим, при выносе правой передней ноги, а следующий вдох - при новом ее выносе, то соотношение ритмов дыхания и движения будет 1:1. Если начало одного вдоха рысака сочетается с выносом правой передней ноги, а начало следующего вдоха - с выносом левой передней (то есть через полтора полных шага), то это соотношение составит 1:1,5.
При совпадении же цикла глубокого дыхания (вдох и выдох) с двумя полными шагами оно будет равно 1:2. В этом случае дыхание является особенно глубоким.
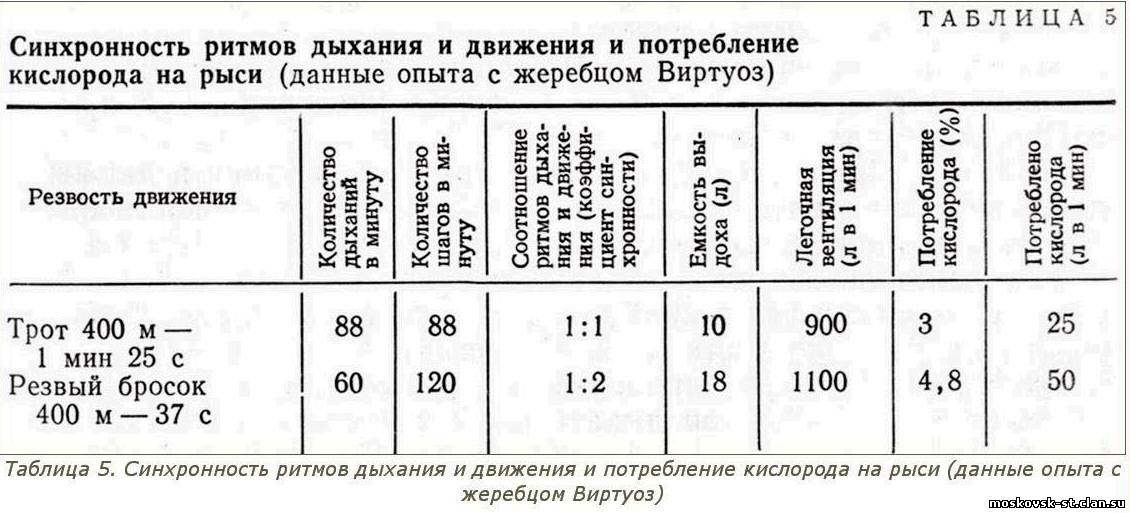
При переходе от синхронности ритмов дыхания и движения в соотношении 1:1 к их синхронности с другим соотношением потребление кислорода намного возрастает. Для наглядности приводим показатели опыта с орловским жеребцом Виртуоз (2.13,2; рожд. 1956 г. от Тульского Пряника и Вазузы) (табл. 5).